内容
- 1 DNA,RNA和蛋白质的合成
-
2 DNA复制
- 2.1 DNA复制错误
-
3 转录
- 3.1 形成的前体信使RNA
- 3.2 RNA剪接
- 3.3 选择性剪接
- 3.4 逆转录
- 4 翻译
- 5 转运RNA
-
6 遗传密码
- 6.1 在使用遗传密码的的运动
- 7 摆动假说
DNA,RNA和蛋白质的合成
遗传物质被存储在DNA的形式在大多数生物体。在人类中,每个细胞的核中含有3×10 9分布在23对染色体的DNA的碱基对,并且每个单元具有遗传物质的两个副本。此统称为人类基因组。人类基因组包含大约30万个基因,其中每一个编码一种蛋白质。
在人类基因组中的DNA的大片被转录,但不为蛋白质编码。这些区域被称为内含子,弥补约95%的基因组。人类基因组的碱基序列,现在已知的精度的一个合理程度,但我们还不知道为什么如此的它是非编码。一些本非编码DNA的控制基因表达,但有多少它的目的尚不清楚。这是一个引人入胜的主题是一定要在未来几年内迅速推进。
在分子生物学中心法则指出,DNA使RNA生成蛋白质(图1)。
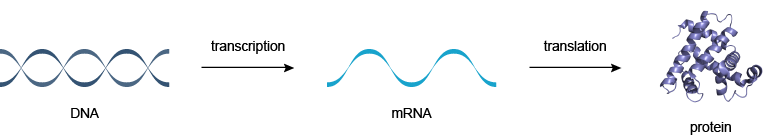
图1 | 分子生物学中心法则:DNA使RNA生成蛋白质
由其中DNA复制的RNA的过程称为转录,并通过其中的RNA被用于产生蛋白质被称为翻译。
DNA复制
每次细胞分裂,各DNA的其双链的分裂成两个单链。每个单链充当互补DNA的新链的模板。其结果是,每一个新的小区都有它自己的完整基因组。这个过程被称为DNA复制。复制是通过在模板链与进入脱氧核苷三磷酸的碱的沃森-克里克配对控制,并通过DNA聚合酶定向。它是一个复杂的过程,特别是在真核细胞中,涉及的酶的阵列。细菌DNA的复制的简化版本中描述如图2。

图2 | 细菌DNA复制简化DNA复制的代表性细菌。
DNA生物合成的收益在5'至3'的方向。这使得无法进行DNA聚合酶同时合成两条链。双螺旋的一部分,必须先放松,这是由导解旋酶。
前导链连续地合成,但相反链被复制在约1000个碱基的短脉冲串,作为随链模板变得可用。由此产生的短链称为冈崎片段(他们的发现者,礼治和冈崎Tsuneko后)。细菌至少有三个不同的DNA聚合酶:POL我,聚合酶II和聚合酶III; 这是波尔三是主要参与链的延伸。奇怪的是,DNA聚合酶不能启动DNA合成从头,但需要用无3'-羟基的短引物。这在由RNA聚合酶(称为DNA的引物酶),其能够在长使用DNA模板和合成一小段RNA的约20个碱基后随链产生。然后聚合酶III可以接管,但最终遭遇在其路径前面合成的短RNA片段中的一个。此时聚合酶I接管,利用其5'-至3'-外切酶活性消化RNA和填充DNA中的间隙,直到达到的DNA的连续拉伸。这使得新合成的DNA的3'端和先前由聚合酶III合成的DNA的5'-末端之间的间隙。该间隙是通过DNA连接酶,使一个5'-磷酸和3'-羟基(间的共价键的酶填充的图3)。DNA复制在先导链起始更为复杂和详细更专门的文本进行了讨论。

图3 | DNA复制DNA聚合酶DNA聚合酶的DNA复制细菌的作用简化表示。
DNA复制错误
DNA复制是不完美的。错误发生在DNA复制,当不正确的碱基掺入生长的DNA链。这导致不匹配的碱基对,或错配。DNA聚合酶有校对活性,和一个DNA修复酶已经进化纠正这些错误。偶尔,错配存活并掺入到基因组中的下一个轮的复制。这些突变可具有没有结果,它们可能会导致生物体的死亡,它们可能会导致遗传性疾病或癌症; 或者他们可以给机体带来竞争优势在其邻国,它通过自然选择导致进化。
转录
转录是DNA拷贝(处理转录)与mRNA,其承载所需蛋白质的合成的信息。转录发生在两大步骤的地方。首先,前信使RNA的形成,与RNA聚合酶的酶的参与。该过程依赖于Watson-Crick碱基配对,和RNA的合成单链是原始DNA序列的反向互补。信使预RNA,然后“编辑”,以产生在这个过程被称为所需的mRNA分子RNA剪接。
信使预RNA形成
转录的机制具有这样的平行DNA复制。作为与DNA复制,必须转录之前发生的双螺旋的部分退绕可以发生,而且是RNA聚合酶的酶催化这一过程。
不像DNA复制,其中两条链被复制,仅一条链被转录。包含基因的链被称为感链,而互补链是反义链。在转录产生的mRNA是有义链的拷贝,但它是被转录的反义链。
核糖核苷三磷酸(国家结核病防治规划)沿反义DNA链调整,与Watson-Crick碱基配对(带U A对)。RNA聚合酶加入核糖在一起形成前体信使RNA分子是反义DNA链的一个区域互补。当RNA聚合酶到达基地三重被读作“停止”信号转录结束。该DNA分子重新风重新形成双螺旋。
中的转录形成的前体信使核糖核酸(橙色)的简化表示。")
图4 | 转录从双链DNA(蓝色)中的转录形成的前体信使核糖核酸(橙色)的简化表示。
RNA剪接
这样形成的前信使RNA的包含未所需蛋白质的合成内含子。信使预RNA被切碎以除去内含子,并在这个过程被称为RNA剪接创建信使RNA(mRNA)(图5)。
剪接。")
图5 | RNA剪接内含子被从前信使RNA,得到信使RNA(mRNA)剪接。
选择性剪接
在选择性剪接,个别外显子或剪接或包括,产生多个不同的可能的mRNA产物。对于不同的蛋白质同种型的每个mRNA产物码; 这些蛋白亚型在其肽序列中,因此它们的生物活性不同。据估计,人基因产物高达60%经历选择性剪接。可变剪接的几种不同的机制是已知的,其中两个在示出图6中。
排除在外,或两个盒外显子可以是互斥的(底部)。")
图6 | 选择性剪接选择性剪接的几种不同的机制存在-的盒外显子既可以包含在或从最终的RNA(顶部)排除在外,或两个盒外显子可以是互斥的(底部)。
选择性剪接有助于蛋白质多样性 - 一个单一的基因转录(RNA),可以有上千种不同的拼接图案,所以会为成千上万的不同蛋白质的代码:从一个相对有限的基因组产生多样化的蛋白质组。拼接是在遗传调节(响应于细胞条件的剪接模式的改变改变蛋白质表达)重要。这也许并不奇怪,不正常的剪接模式可能导致疾病,包括癌症。
逆转录
在反转录,RNA被“反转录”到的DNA。这个过程中,用逆转录酶催化的,允许逆转录病毒,包括人类免疫缺陷病毒(HIV),为使用RNA作为遗传物质。逆转录酶人还发现在生物技术的应用,使科学家能够RNA转变成DNA为技术如PCR。
翻译
形成转录mRNA的运出细胞核,进入细胞质,核糖体(细胞的蛋白质合成工厂)。这里,它指导蛋白质的合成。信使RNA不直接参与蛋白质的合成-转运RNA(tRNA的)需要这一点。通过该mRNA的指导蛋白质合成与tRNA的协助下,对过程称为翻译。
核糖体是RNA和蛋白质分子的一个非常大的复合物。的mRNA(三重峰)的各三碱基拉伸被称为一个密码子,和一个密码子包含特定氨基酸的信息。作为mRNA的穿过核糖体,每个密码子与交互反密码通过Watson-Crick碱基配对的特定转移RNA(tRNA的)分子。此的tRNA分子携带在其3'末端,其被结合到生长的蛋白质链的氨基酸。tRNA的然后从核糖体排出。图7示出了参与蛋白质合成的步骤。
和(b)的tRNA分子结合到核糖体的两个结合位点,并通过氢键与mRNA; (c)该两个氨基酸之间的肽键的形式,使二肽,而tRNA的分子是左不带电荷的; (d)该不带电荷的tRNA分子离开核糖体,而核糖体移动一个密码子在正确的(二肽是由一个结合位点易位到其他); (五)其他的tRNA分子结合; (六)在两个氨基酸之间的肽键的形式使三肽; (七)不带电荷的tRNA分子离开核糖体。")
图7 | 翻译(一)和(b)的tRNA分子结合到核糖体的两个结合位点,并通过氢键与mRNA; (c)该两个氨基酸之间的肽键的形式,使二肽,而tRNA的分子是左不带电荷的; (d)该不带电荷的tRNA分子离开核糖体,而核糖体移动一个密码子在正确的(二肽是由一个结合位点易位到其他); (五)其他的tRNA分子结合; (六)在两个氨基酸之间的肽键的形式使三肽;(七)不带电荷的tRNA分子离开核糖体。
转移RNA

图8 | 的tRNA(转运RNA)的二维结构。在一些的tRNA的DHU臂只有三个碱基对。
每种氨基酸都有其特殊的tRNA(或一组的tRNA)。例如,所述的tRNA为苯丙氨酸(tRNAPhe)是从不同的组氨酸(tRNAHis)。各氨基酸通过3'-OH基团连接到其的tRNA以形成与α氨基生长蛋白链的末端氨基酸的反应的酯,以形成蛋白质中一个新的酰胺键(肽键)合成(图9)。酯与胺的反应通常是有利的,但反应速度在核糖体大大增加。

图9 | 蛋白质合成与带电的tRNA的3'末端的生长的多肽链的反应。的氨基酸是从的tRNA分子到蛋白质转移。
每个转移RNA分子具有被酶氨酰基tRNA合成酶,它增加了正确的氨基酸的不带电荷的tRNA的3'端识别的良好定义的三级结构。修饰的核苷的存在是在稳定的tRNA结构重要。一些这些修改的示于图10。

图10 | 在tRNA的修饰的碱基的一些发现的tRNA的修饰碱基的结构。
遗传密码
遗传密码几乎是普遍的。它是由核酸在所有生物体的遗传信息传递的基础。有在RNA(A,G,C和U)四种碱基,所以有64种可能的三联码(4 3 = 64)。理论上仅需要22码:每个20种天然存在的氨基酸,增加一个起始密码子和终止密码子(以指示蛋白质序列的开始和结束)。许多氨基酸具有几个码(简并性),因此,使用所有64种可能的三联体编码。例如精氨酸和服务各有6个密码子,而色氨酸和蛋氨酸只有一个。没有两个氨基酸具有相同的代码,但氨基酸的侧链具有相似的物理或化学性质往往有相似的密码子的序列,例如苯丙氨酸,亮氨酸,异亮氨酸,缬氨酸都是疏水性的,和Asp和Glu的侧链两个羧酸(参见图11)。这意味着,如果在翻译过程中选择了不正确的tRNA(在密码子反密码接口由于单一碱的错配)的misincorporated氨基酸可能具有类似的性质,以预定的tRNA分子。虽然所得到的蛋白质将具有一个不正确的氨基酸它代表的是功能性的可能性高。生物显示“密码子偏好”,并使用特定的密码子的特定氨基酸比别人多。例如,在人类中的密码子使用是与在细菌不同; 它有时可能难以在细菌中表达人蛋白因为相关的tRNA可能存在在太低的浓度。
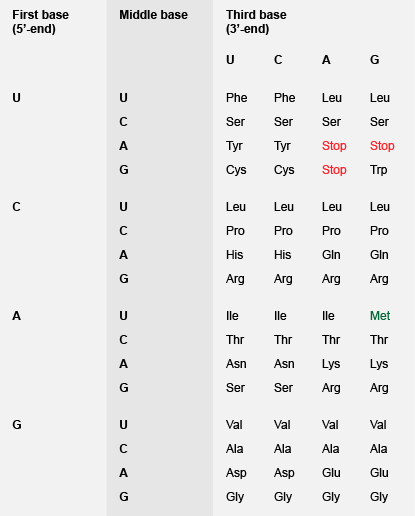
图11 | 遗传代码-三重密码子分配为20个氨基酸。以及编码蛋氨酸,AUG被用作起始密码子,起始蛋白质的生物合成
在使用遗传密码的一个运动
基因组DNA(链A,编码链)的一条链包含下列序列读取从5'到3':
TCGT
这股将形成以下复式:
5'-TCGTA-3'
3'-AGCAT-5'
因此在DNA(链B)的写入5'-至3'-另一条链碱基序列是
TCGAG
基地的mRNA从书面到5'3'的DNA链转录的序列
UCGAG
由上述的mRNA编码的氨基酸序列是
丝氨酸 - 丝氨酸 - 丝氨酸 - 精氨酸 - 停止
然而,如果DNA链B是编码链mRNA序列将是:
UCG
和氨基酸序列将是:
丝氨酸 - 丝氨酸 - 苏氨酸 - 精氨酸 - 丝氨酸 - 丝氨酸 - 甘氨酸 - 半胱氨酸 - 丝氨酸
摆动假说
所有特定氨基酸可用密码子的靠近检查发现的变化是在第三位置最大(例如,丙氨酸的密码子是GCU,GCC,GCA和GCG)。克里克和布伦纳提出单一的tRNA分子可以识别与在由于非Watson-Crick碱基对的形成与在密码子反密码相互作用第三基3'-端不同碱基的密码子。这些非标准碱基对是由形状不同·U和·G·C和术语摆动假说表明有一定程度的柔性或“摆动”是允许在核糖体这个位置。不是所有的组合都是可能的; 的“允许”的配对的例子示于图12。
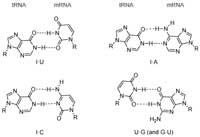
图12 | 摆动碱基对的结构中发现的RNA
DNA碱基,以形成摆动碱基对以及Watson-Crick碱基对的能力可导致碱基对错配的DNA复制过程中发生的。如果不被修复的DNA修复酶,这些错配可导致遗传性疾病和癌症。