内容
2'-脱氧核糖核酸(DNA)的
DNA(2'-脱氧核糖核酸)是遗传信息几乎所有生命系统的分子店。它是已知的作为单体的由大量的聚合物分子的核苷酸。每个核苷酸组成的杂环基,戊糖(2'-脱氧的ð -ribofuranose),和磷酸基。有DNA中四种杂环碱基:腺嘌呤(A),鸟嘌呤(G),胞嘧啶(C)和胸腺嘧啶(T)。其结构和编号系统示于图1。

图1 | DNA的杂环碱基的化学结构
腺嘌呤和鸟嘌呤是嘌呤和胞嘧啶和胸腺嘧啶是嘧啶(图2)。

图2 | 的嘌呤和嘧啶杂环系统的结构
所述脱氧核糖示于图3中作为一个无糖它能。变旋在某些条件下,采用呋喃糖,无环和吡喃糖形式; 但在DNA中它固定为呋喃。

图3 | 2'-脱氧-D-核糖的变旋允许呋喃糖,无环和吡喃糖形式之间的互
磷酸基团可以在根据用于分解的DNA,以产生核苷酸的方法中的糖的5'或3'位被发现。磷酸盐的去除产生了一个核苷。该杂环基是与糖的1'-位上。一脱氧和脱氧核苷酸的化学结构示于图4。

图4 | 一脱氧和脱氧核苷酸的结构
债券加入脱氧核糖的1'碳的杂环基地是ñ糖苷键。有关此键旋转引起顺和反构象()。这个键的旋转受到限制和反构象的普遍青睐,部分原因是空间的理由。
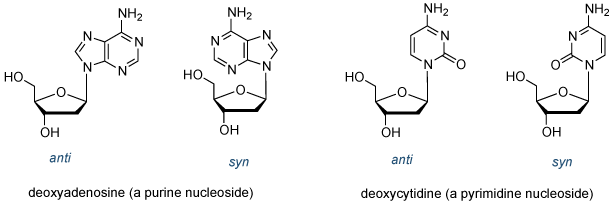
图5 | 的结构顺式和反核苷构
在DNA双链体的核苷采用反构象(也有极少数例外情况,其中之一是鸟苷酸,其中,所述鸟嘌呤碱基采用顺约糖苷键构)。
所述脱氧核糖的1'-位是端基异构体中心。如果附着到1'-碳取代基位于同一面上的糖环作为5'-羟基,它被称为β型异头物; 如果取代基是在糖环的相对侧它是α端基异构体(图6)。所有在DNA中的核苷的是将β构型。

图6 | 核苷α-的结构和β端基异构体
核糖核酸(RNA)
一些生物体,例如逆转录病毒,用核糖核酸(RNA),而不是DNA作为其遗传信息存储。RNA的化学性质非常相似的DNA,但有两个重要的区别:- 的RNA具有连接到糖的2'位上的羟基基团,和
- 嘧啶尿嘧啶(U; 图7)中的RNA代替胸腺嘧啶。
RNA的关键生物作用是作为一个信使:它在DNA中(读遗传密码转录)并将其传送到核糖体,在那里被解码成蛋白质(序列翻译)。

图7 | 尿嘧啶和胸腺嘧啶核苷结构尿苷的RNA发生; 胸苷的DNA。
寡核苷酸
DNA或RNA的核苷酸(二聚体)是由一个核苷酸的5'-磷酸基团共价连接到另一个的3'-羟基基团,以形成一形成磷酸二酯键。当几个这样的键被制成,以及天然存在的核酸是这样的直链,高分子量分子形成的寡核苷酸(寡聚物)。在生理pH(7.4)每个磷酸二酯基团的存在作为阴离子(因此术语核酸),因此核酸高度带电聚阴离子分子(图8)。
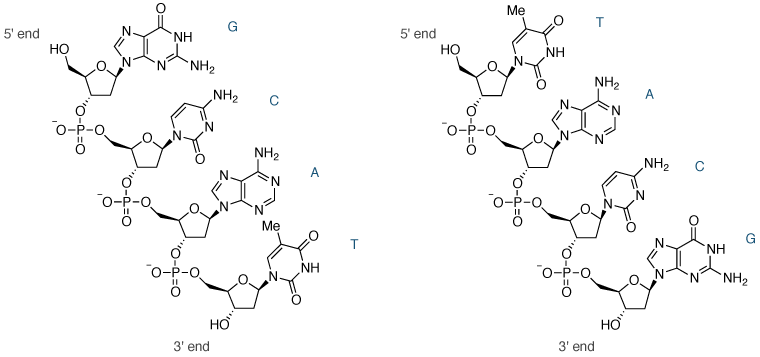
图8 | 寡核苷酸dGCAT(左)和dTACG的化学结构(右)
核酸链的一端具有5'-羟基(伯羟基),另一端有一个3'-羟基(仲羟基)。因此,核酸链具有方向性。在四核苷酸图8一个具有序列5'-GCAT-3',并在四核苷酸图8B具有序列5'-TACG-3'。按照惯例,使用前缀5'-和3'-未写入,和核酸序列被写入的5'到3'的方向。寡核苷酸GCAT和TACG是具有不同的化学和生物物理特性不同的分子。
核酸双链体
DNA的单链的化学结构使得不能洞察其生物功能的遗传信息的载体。然而,当詹姆斯·沃森和弗朗西斯·克里克显示,1953年采用DNA双链结构(双面),机制DNA复制(复制)变得明显。双螺旋结构,主要由X射线衍射纤维鉴定数据和查戈夫的规则(由罗莎琳德·富兰克林和威尔金斯收购)。欧文查戈夫发现在DNA中的腺嘌呤的摩尔量总是等于胸腺嘧啶和相同的是鸟嘌呤和胞嘧啶(即的G摩尔= C的摩尔数)真。沃森和克里克能够通过建立模型以显示DNA的两条链通过在相反链单个碱基之间的氢键保持在一起来解释这一点。嘌呤碱基A总是与嘧啶T和嘌呤ģ总是成对与嘧啶-C(对图9)。

图9 | 在氢键·T和C·G的Watson-Crick碱基对
毫不奇怪的A·T和G·C被称为Watson-Crick碱基对。它们是伪对称并且如果A·T碱基对铺设在任何其他碱基对(T·A,G·C或C·G)的磷酸二酯主链落在彼此的顶部(图10和图11)。因此,所有四个碱基对正好贴合了双螺旋结构。
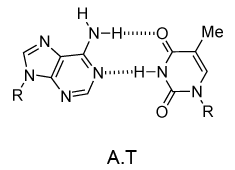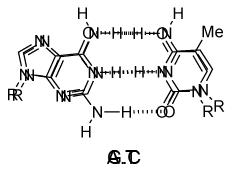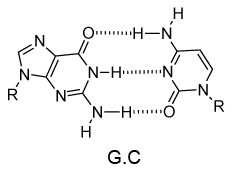
图10 | ·G·C和A的动画叠加·T碱基对

图11 | 碱基对重叠的T·碱基对覆盖的A·T碱基对,而对G·C碱基对覆盖在A·T碱基对的结构。
DNA的一条链的序列精确定义了其他的序列; 两条链被说成是互补的,并且有时被称为反向互补彼此的。两条链是反平行的,与相邻的其他的3'端的一条链的5'末端。两个链彼此盘绕以形成右旋双螺旋,以形成外部亲水性主链中的中心疏水碱基对和糖和带负电荷的磷酸盐。术语“右旋”表示在面向观察者的分子的前端的骨干从到左下方右上方向下倾斜。平面杂环碱堆叠在另一个上并沿所述螺旋轴连续的碱基对之间的间隔大约是0.34纳米。一个螺旋圈(一个完整的360°旋转的双螺旋的)被重复,每10至11个碱基对。所述双链体的稳定性是从两者得到的碱基堆积和氢键。双螺旋的DNA,B-DNA(的主要形式图12,中间),具有很宽的大沟和周围沿该分子的整个长度的螺旋运转的窄的小沟。蛋白质在这些凹槽(主要在大沟)和一些小分子药物(如纺锤菌素,偏端霉素)结合的DNA在小沟(见互动核酸药物相互作用)。
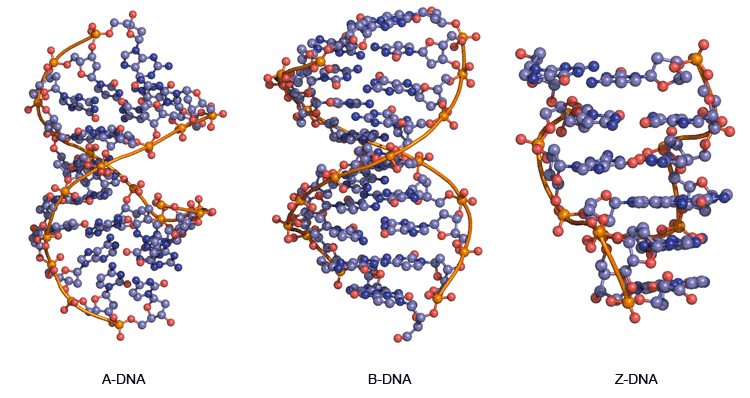
图12 | A-DNA的三维结构,B-DNA和Z-DNA
RNA也可以使用相同的碱基配对规则形成右旋双工(A·U和·G·C)中,但将RNA双链体具有独特的形状(A型),其中大沟深,小沟很浅。下(低湿度的DNA条件下也能采用的A型图12,左),以及有许多DNA和RNA的其它构象,其中大部分是在A-和B-形式微妙的变化。
一个显着不同的DNA构象,Z-DNA,(图12,右)从交替序列dCGCGCG,其自发地形成在含水缓冲液中的双链体的化学合成的DNA链的X射线晶体结构确定。Z-DNA是左撇子,并具有一个二核苷酸重复单元,所以主链不顺畅,但似乎“之字形”。目前尚不清楚Z-DNA是否具有生物相关性。
确定核酸结构技术
核磁共振(NMR)
核磁共振是高度发达的和强大的光谱技术是在核酸的结构,热力学和动力学性能的调查有价值。该技术可以用于研究DNA双链体,三螺旋,四链,发夹环,RNA双链和其他二级和三级RNA结构。首先,1 H-NMR与水抑制水性缓冲液使用。31 31 P NMR也是有用研究核酸磷酸二酯骨架的环境。核磁共振是在当地研究核酸结构,即碱基对和三重峰,核酸-药物相互作用和核酸-蛋白质相互作用的性质最有用的。这是给全球性的信息(如A与B螺旋,DNA双螺旋弯曲),因为当地的互动必须作为全球性的指标那么强大。核磁共振应始终与其它物理技术,如圆二色性一起使用(指示构象中,A,B,Z),的双链体,三螺旋和四链(热力学信息)紫外线熔化,X射线衍射(高分辨率结构分析)和荧光共振能量转移(距离测量)。碱和糖质子典型核磁共振化学位移列于图13和图14。
。")
图13 | 在核酸碱基的1 H NMR谱中观察到的关键残基的NMR化学位移从Wijmenga和van布伦,数据在核磁共振谱进展 32,287-387(1998)。
。")
图14 | 在脱氧核糖的1H NMR谱观察核酸的关键残基的NMR化学位移从Wijmenga和van Buuren的,数据在核磁共振谱进展 32,287-387(1998)。
结晶和X-射线衍射
在进步固相寡核苷酸合成和纯化已允许大量寡核苷酸的待结晶和其分子结构解决。这提供了B和A形式的DNA的两个上重要的信息,在确认从早期纤维衍射研究作出的预测,并提供关于核酸结构的序列依赖性的高分辨率信息。单晶,例如在所示的X射线衍射图15提供大分子结构的精确图象与超过其它技术的分辨率。
当晶体被放置在X射线源,将产生的衍射图案,从其中,一些复杂的数学的帮助下,该单位电池的立体图像和分子本能够生成的路径。X射线晶体需要高度有序晶体,并获得核酸合适的晶体一般是该方法的最慢的部分。的寡核苷酸的高度浓缩的溶液需同时提供晶体形成热力学驱动力。溶解度极限通过使用外部脱水剂从所述寡核苷酸溶液逐渐除去水,或扩散沉淀如异丙醇到溶液中超过。已通过X射线晶体学获得的A,B和Z-DNA的高分辨率结构和技术已经被用来研究在原子细节DNA的药物复合物。在大多数情况下,该结构的分辨率不超出2埃(0.2纳米),但是这是足以提供杂环碱基和糖 - 磷酸酯主链的一个合理的清晰画面。

图15 | 的DNA晶体的DNA寡核苷酸的晶体的照片。